Methodology of Leaf Anatomical Structure and
Geographic Environmental Dataset of Caryopteris mongholica from Seven
Regions in North and Northwest China
Ji, R. X. Yu,
X. Chang, Y. Shen, C. Bai, X. Q. Xia, X. L. Yin, W. L. Liu, C.*
National Engineering Laboratory of Tree Breeding, College
of Biological Sciences and Technology, Beijing Forestry University, Beijing
100083, China
Abstract: The genetic variation caused by long-term adaptation to the
environment significantly affects plant growth and development. Leaves are the
most important and sensitive organs in responding to environmental changes.
Therefore, an understanding of the adaptive variation in leaf anatomical
structures in different environments is the basis for exploring plant
adaptation to the environment. Common garden experiments eliminate the influence of environmental gradients and are an effective approach
to study the effects of genetic and environmental factors on plant structures. In this study, a common
garden experiment was used to analyze the anatomical structures of Caryopteris
mongholica leaves from seven different areas in north and northwest China,
including Abaga Banner, Alxa Left Banner, Dongwu Banner, Mengxi, and Liangcheng
in Nei Mongol (Inner Mongolia), Shenmu in Shaanxi province, and Kangbao in
Hebei province. Conventional paraffin sectioning was used to analyze
structures, and multiple comparisons, correlation analyses, and a general
linear model (GLM) were used to identify the factors driving differences. The
genetic variation driven by climate was one of the major factors that caused
the differences in leaf anatomical structures from the different areas. The
dataset includes the following: (1) geographical location data for the
collection sites; (2) C. mongholica leaf cross sections; and (3) analysis of the relations
between C. mongholica leaf anatomical structures and the geographic
environmental factors that influence
them. The dataset is archived in .shp, .kmz, .xls, .jpg, and .doc
formats and consists of 14 data files, with a data size of 5.84 MB.
Keywords: Caryopteris mongholica; common garden
experiment; leaf anatomical structures
Dataset Availability Statement:
The dataset supporting this paper was published and is accessible
through the Digital
Journal of Global Change Data Repository at:
https://doi.org/10.3974/geodb.2021.01.03.V1.
1 Introduction
Plants are subject to the
long-term effects of environmental factors such as temperature, water, and
light and in the process of evolution, gradually develop phenotypic and genetic
characteristics adapted to the environment. Even within the same species,
long-term growth in different geographic environments can produce different
degrees of gene differentiation that lead to specific regional characteristics
adapted to local geographic environments[1–4]. The leaf is the plant organ
with the largest area exposed to the environment, and its external
morphological characteristics and internal anatomical structure can best
reflect the adaptive evolution of plants to environmental factors[5–8]. Therefore,
analyzing the anatomical structure of leaves is important in studying the adaptation
strategies of plants in extreme environments.
Common garden experiments are
used to minimize the effects of site conditions[9,10] and show that differences in
leaf anatomical structures from different regions are due to the genetic
variation in plants caused by environmental differences in the original regions.
This experiment differs from previous ones on the characteristics of leaf anatomical
structure, because the effects of both environmental factors, including climate
indices and geographic location, and genetic factors were evaluated. Common
garden experiments can provide important insights into local adaptability[11] and a better understanding of the
responses and adaptation mechanisms of plants from different geographical
regions to global changes. At present, few studies have used this method to
analyze the differences in leaf anatomical structures from different regions
and the factors that influence changes in those structures. Therefore, Caryopteris mongholica
from seven regions in north and northwest China were planted under the same
environmental conditions in a common garden experiment to exclude the effects
of environmental factors. To explore the effect of locality on the plant,
paraffin sectioning was used to study leaf anatomical structures. With an
understanding of adaptive evolution according to regional environments, a
theoretical basis is provided for protection of germplasm resources, genetic
improvement, and exploration of the mechanisms by which differences in plant
genotypes are driven by environmental variation.
2 Metadata of the Dataset
The metadata
of Leaf anatomical structure and geographic environment data of Caryopteris Mongholica from 7 provenances[12] are summarized in Table 1, including the dataset full name, short name,
authors, year of the dataset, temporal resolution, data format, data size, data
files, data publisher, and data sharing policy, etc.
3 Data Development Methods
3.1 Study Area
The common
garden experiment was conducted in the nursery of Beijing Forestry University (40.01??N, 116.34??E), China. The
nursery is in the warm temperate zone with a semihumid continental monsoon
climate. The mean annual temperature is 11.8 ??C, the annual maximum temperature range is 37.5–42.6 ??C,
the annual minimum temperature range is –19.5 to –14.8 ??C, and the mean annual
precipitation is 500 to 650 mm[14]. Most precipitation occurs in the
summer, accounting for 70% of the annual total, and severe drought is typical
in spring. The research site was covered with a plastic shed
overnight and when it rained to prevent the influence of additional water.
3.2 Plant Materials
One-year-old C. mongholica cuttings, approximately
20-cm stems, with good growth were collected. They were wrapped in damp
cloth and immediately brought to Beijing. The cuttings were collected
from Abaga Banner, Alxa Left Banner, Dongwu Banner, Mengxi, and Liangcheng in
Nei Mongol (Inner Mongolia), Shenmu in Shaanxi province, and Kangbao in Hebei
province in late March 2016. The cuttings from the different regions (Table 2)
were treated with 0.1% rooting powder ABT1 (Research Institute of Forestry,
Chinese Academy of Forestry) and then planted in flowerpots (450 mm ?? 450 mm). The
soil was a sandy loam, and the volume ratio of peat soil, loam soil, and sand
was 4:4:3, which is similar to the composition of the field soil in which C.
mongholica was collected. All materials were planted in the same soil mix.
Table 1 Metadata summary of the Leaf
anatomical structure and geographic environment data of Caryopteris Mongholica from 7 provenances
|
Items
|
Description
|
|
Dataset full name
|
Leaf
anatomical structure and geographic environment data of Caryopteris Mongholica from 7 provenances
|
|
Dataset short name
|
LeafAnatomicalStructure_CaryopterisMongholica
|
|
Authors
|
Ji, R. X. AAE-6059-2021,
Beijing Forestry University, bljrx@bjfu.edu.cn
Yu, X. AAE-6050-2021, Beijing
Forestry University, yuxiao@bjfu.edu.cn
Chang, Y., Beijing Forestry
University, 1034530229@qq.com
Shen, C. 0000-0001-5037-4922,
Beijing Forestry University, shenchaonow@bjfu.edu.cn
Bai, X. Q., Beijing Forestry
University, baixueqia88@qq.com
Xia, X. L., Beijing Forestry
University, xiaxl@bjfu.edu.cn
Yin, W. L., Beijing Forestry
University, yinwl@bjfu.edu.cn
Liu, C. AAE-6091-2021, Beijing
Forestry University, liuchao1306@bjfu.edu.cn
|
|
Geographical region
|
China 38.88??N–45.73??N, 105.72??E–116.79??E
|
Year
|
2007–2017
|
|
Temporal resolution
|
Day
|
|
|
|
|
|
Data format
|
.shp, .kmz, .xls, .jpg and
.doc
|
Data size
|
5.84 MB
|
|
Data files
|
(1) Geographical location data of collection sites
(2) Leaf cross section of Caryopteris
mongholica
(3) Analysis data of leaf anatomical structure and geographical environment
influencing factors of Caryopteris mongholica:
1) Summary of
general linear models for the effect of climate variations in different regions
on individual leaf anatomical characters of Caryopteris mongholica
2) Correlation
analyses data of Caryopteris mongholica leaf
anatomical characters
3) Leaf anatomical
parameters and basic
conditions of Caryopteris mongholica regions
|
|
Foundations
|
Ministry of Ecological
Environment of P. R. China (2017ZX07101002); National Natural Sciences
Foundation of China (32071734, 31770649, 31600484)
|
|
Data Computing Environment
|
ArcGIS 10.2, Excel, R 3.2.2
|
|
Data publisher
|
Global Change Research Data
publishing & Repository, http://www.geodoi.ac.cn
|
|
Address
|
No.11A, Datun Road, Chaoyang
District, Beijing 100101, China
|
|
Data sharing policy
|
Data from the Global Change
Research Data Publishing & Repository includes metadata, datasets (in the Digital Journal of Global Change Data Repository), and
publications (in the Journal of Global Change Data & Discovery). Data
sharing policy includes: (1) Data are openly available
and can be free downloaded via the Internet; (2) End users are encouraged to
use Data subject to citation; (3) Users, who are by definition
also value-added service providers, are welcome to redistribute Data subject
to written permission from the GCdataPR Editorial Office and the issuance of
a Data redistribution license; and (4) If Data are
used to compile new datasets, the ??ten per cent principal?? should be followed
such that Data records utilized should not surpass 10% of the
new dataset contents, while sources should be clearly noted in suitable
places in the new dataset[13]
|
|
Communication and
searchable system
|
DOI, DCI, CSCD, WDS/ISC,
GEOSS, China GEOSS, Crossref
|
|
|
|
|
|
|
From three to five
cuttings with similar growth from each site, three mature leaves in the middle
of each branch were sampled. A 1 cm ?? 1 cm square (approximately) was cut from
the middle of each leaf that included the main vein (Figure 1). The pieces were
fixed in FAA (formaldehyde:acetic acid:70% ethanol = 1:1:18) and taken to the
laboratory for determination of leaf anatomical structures.
Table
2 Locations
and climate indices of Caryopteris mongholica collection sites in this
study
|
Collection sites
|
Latitude (??N)
|
Longitude (??E)
|
Altitude (m)
|
MAP (mm)
|
MAT (??C)
|
GSP (mm)
|
GST (??C)
|
TCM (??C )
|
PE (mm)
|
|
Abaga Banner, Nei Mongol
|
43.90
|
115.35
|
1,177
|
224.03
|
2.9
|
434.71
|
17.74
|
–19.85
|
588.04
|
|
Alxa Left Banner, Nei Mongol
|
38.88
|
105.72
|
1,670
|
162.54
|
9.90
|
315.52
|
21.05
|
–7.51
|
712.81
|
|
Dongwu Banner, Nei Mongol
|
45.73
|
116.79
|
1,017
|
200.72
|
2.46
|
415.46
|
17.76
|
–20.52
|
531.06
|
|
Mengxi, Nei Mongol
|
40.08
|
106.92
|
1,193
|
118.00
|
8.92
|
243.64
|
20.95
|
–9.58
|
702.17
|
|
Liangcheng, Nei Mongol
|
40.66
|
112.30
|
1,429
|
313.75
|
5.18
|
611.46
|
17.13
|
–13.17
|
580.80
|
|
Shenmu, Shaanxi
|
39.29
|
110.33
|
1,209
|
369.79
|
9.78
|
729.85
|
20.85
|
–7.64
|
706.91
|
|
|
41.99
|
114.85
|
1,590
|
279.72
|
3.82
|
556.53
|
16.78
|
–15.22
|
544.06
|
Note: MAP,
mean annual precipitation; MAT, mean annual temperature; GSP, growth season precipitation; GST, growth
season temperature; TCM, temperature of coldest month; PE, potential evaporation.
|

Figure 1 Leaf section
cut for determination of anatomical structures
|
3.3
Methods and Algorithm Principle
Conventional paraffin sectioning was used to analyze the
leaf structures[15]. The leaf squares were fixed for more than 24 h and then
dehydrated in a four-step gradient of 70%, 85%, 95%, and 100% alcohol. The
covers were sealed to prevent water entering the air space, and xylene was used
to increase the transparency of leaf material. The leaf squares were embedded
in liquid paraffin (paraffin melting point =
56–57 ??C). After the paraffin solidified, blocks
were sliced into sections 8 to 10 µm thick. The sections were stained with
safranin-fast green and mounted in neutral gum[16]. A Leica DM2500 microscope with Leica LAS AF software was
used to observe and analyze the sections (Leica, Wezlar, Germany).
Image J software was used to measure the thickness of the cuticle, the upper and lower
epidermal cells (UEC, LEC), the upper and lower palisade tissues(UPT,
LPT), and the leaf thickness(LT). The tightness of the leaf and
the total thickness of the palisade tissue(PT) were calculated[17]. For
each index, the measurement was repeated five times.
Leaf
anatomical indices:
Tight = PT/LT
?? 100%
PT = UPT
+ LPT
Plasticity indices of leaf anatomical
structures:
Coefficient of variation = Standard
deviation / Arithmetic mean
Plasticity index = (Maximum – Minimum) / Maximum
According to the longitude and latitude of the
different C. mongholica collection sites, the monthly average climate data (2007–2016) of each sample site were obtained from the
WorldCLIM global high-resolution climate database of ArcGIS 10.2 and used to
calculate the climate indices. The climate indices included mean annual
precipitation (MAP), growth season precipitation
(GSP), annual range of precipitation
(ARP), mean annual temperature (MAT), growth season temperature (GST), annual range of temperature (ART), and potential evaporation (PE).
3.4 Data Analysis
A
multiple comparisons method was used to compare differences in the anatomical
structures of C. mongholica. Pearson correlations were used to analyze
the relations between leaf anatomical structures and environmental factors.
Combined with a general
linear model (GLM) ANOVA to analyze the effects of collection-site climate on
the anatomical structures[19]. Based on the correlations between
collection-site environmental factors and leaf anatomical structures, MAP,
MAT, PE, Alt, and
Site were selected as environmental factors that were included in the
model.
Data
analyses were conducted in Excel and R 3.2.2.
3.5 Data Development
Flowchart
The
following steps were used to produce the dataset (Figure 2): Caryopteris mongholica
was collected in seven different sites based on previous data collection and
field investigations. The C. mongholica were planted in the nursery of
Beijing Forestry University in a common garden experiment. After 18 months,
samples were cut from C. mongholica leaves and fixed. Leaf anatomical
structures were measured. Correlations were conducted between the anatomical
structures themselves and between geographic environmental factors and
anatomical structures. The GLM was used to verify the significant factors of
influence. Thus, leaf anatomical structure and geographic environmental
datasets of C. mongholica collected from seven sites were formed.
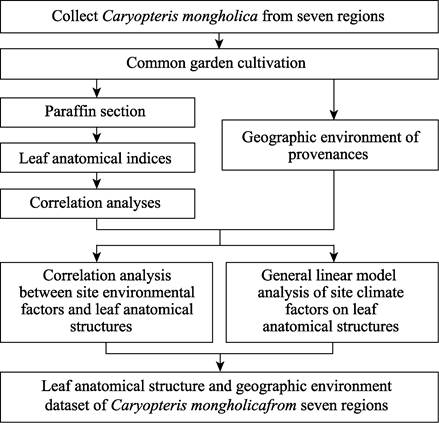
Figure 2 Technical flowchart of the
analysis of leaf anatomical structures of Caryopteris mongholica and the environmental factors of influence
4 Data Results and Verification
4.1 Dataset composition
The dataset is composed of four
files. The data files include geographical location data of the
collection sites; 2007 to 2016 climate indices; leaf cross sections and parameters; and GLM
analysis of regional climate factors and leaf anatomical structures. The dataset is stored in .shp,
.kmz, .xls, .jpg, and .doc formats.
4.2 Data Results
Caryopteris mongholica from the seven collection sites had typical isolateral
leaves, and the thickness of leaves was between 192.34 and 270.30 µm (Table 3). The internal structure of leaves
was divided into the epidermis, mesophyll, and veins (Figure 3).
Table 3 Leaf anatomical indices of
Caryopteris mongholica
|
Anatomical indices
|
Number of
observations
|
Average
|
Standard deviation
|
95% Confidence
interval of mean
|
Maximum
|
Minimum
|
Coefficient of variation
|
Plasticity index
|
|
Cuticle (µm)
|
70
|
6.87
|
1.66
|
6.05–7.70
|
11.92
|
5.07
|
0.24
|
0.57
|
|
UECµm)
|
70
|
18.53
|
5.37
|
15.86–21.20
|
26.46
|
9.87
|
0.29
|
0.63
|
|
LEC µm)
|
70
|
11.14
|
2.32
|
9.99–12.30
|
17.64
|
8.74
|
0.21
|
0.50
|
|
UPT µm)
|
70
|
106.35
|
17.94
|
97.43–115.27
|
146.64
|
89.24
|
0.17
|
0.39
|
|
LPT µm)
|
70
|
76.17
|
13.44
|
69.48–82.85
|
109.42
|
59.57
|
0.18
|
0.46
|
|
PT µm)
|
70
|
182.52
|
29.16
|
168.02–197.02
|
243.12
|
156.05
|
0.16
|
0.36
|
|
LT µm)
|
70
|
220.02
|
25.71
|
207.24–232.80
|
270.30
|
192.34
|
0.12
|
0.29
|
|
Tight (%)
|
70
|
0.83
|
0.04
|
0.80–0.85
|
0.90
|
0.74
|
0.05
|
0.18
|
Note: UEC, upper epidermal cell; LEC, lower epidermal cell; UPT, upper palisade tissue; LPT, lower palisade tissue; PT, palisade tissue; LT, leaf thickness.
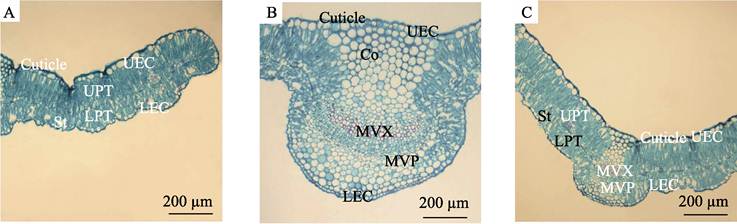
Figure 3 Leaf cross sections of Caryopteris
mongholica (A. Mesophyll
structure; B. Main vein structure; C. Mesophyll and main vein structure)
(Note: Co, collenchyma; LEC,
lower epidermal cell; LPT,
lower palisade tissue; MVP,
main vein phloem; MVX, main
vein xylem; St, stomatal
chamber; UEC, upper epidermal
cell; UPT, upper palisade
tissue)
Most of the leaf anatomical structures were
correlated with other structures, with UPT, LPT,
PT, LT, and Tight highly significantly positively
correlated with other structures (P < 0.01, Table 4).
4.3 Dataset Verification
Leaf
anatomical structures were significantly correlated with one or more of geographical
location (latitude, longitude), temperature, and precipitation (Table 5).
Longitude
and latitude were highly significantly positively correlated with UEC
but were significantly negatively correlated with UPT, LPT, PT,
LT, and Tight (P < 0.05). On the geographical gradient,
UEC gradually increased from west to east and south to north, whereas UPT,
LPT, PT, LT, and Tight gradually decreased.
Table 4 Pearson coefficients of
correlation between leaf anatomical structures of Caryopteris mongholica
|
Leaf characters
|
Cuticle µm)
|
UEC µm)
|
LEC µm)
|
UPT µm)
|
LPT µm)
|
PT µm)
|
LT µm)
|
Tight (%)
|
|
Cuticle µm)
|
|
|
|
|
|
|
|
|
|
UEC µm)
|
0.201
|
|
|
|
|
|
|
|
|
LEC µm)
|
0.590**
|
0.107
|
|
|
|
|
|
|
|
UPT µm)
|
–0.246
|
–0.355
|
–0.203
|
|
|
|
|
|
|
LPT µm)
|
–0.434
|
–0.545*
|
–0.436
|
0.721**
|
|
|
|
|
|
PT µm)
|
–0.351
|
–0.470*
|
–0.326
|
0.948**
|
0.904**
|
|
|
|
|
LT µm)
|
–0.260
|
–0.268
|
–0.259
|
0.945**
|
0.825**
|
0.962**
|
|
|
|
Tight (%)
|
–0.432
|
–0.770*
|
–0.391
|
0.683**
|
0.806**
|
0.792**
|
0.595**
|
|
*P <
0.05; **P < 0.01.
Note: UEC, upper epidermal cell; LEC, lower epidermal cell; UPT, upper palisade tissue; LPT, lower palisade tissue; PT, palisade tissue; LT, leaf thickness.
Leaf UPT, LPT, PT, LT,
and Tight were all significantly positively correlated with MAT (P
< 0.05), whereas UEC was
significantly negatively correlated with MAT and GST (P
< 0.05) and significantly positively correlated with ART (P
< 0.05). As temperature increased, UPT, LPT, PT, LT,
and Tight increased and UEC decreased.
The
precipitation indices MAP, GSP, and ARP were only
significantly negatively correlated with UPT (P
< 0.01), LT (P < 0.05), and PT (P < 0.05).
Table 5 Pearson coefficients of
correlation between environmental factors at collection sites and leaf
anatomical structures of Caryopteris mongholica
|
Environmental factors
|
Cuticle µm)
|
UEC µm)
|
LEC µm)
|
UPT µm)
|
LPT µm)
|
PT µm)
|
LT µm)
|
Tight %)
|
|
Latitude (??N)
|
0.328
|
0.638**
|
0.451
|
–0.526*
|
–0.694**
|
–0.643**
|
–0.529*
|
–0.724**
|
|
Longitude (??E)
|
0.437
|
0.704**
|
0.366
|
–0.787**
|
–0.801**
|
–0.854**
|
–0.730**
|
–0.886**
|
|
Altitude (m)
|
–0.124
|
–0.034
|
–0.358
|
0.547*
|
0.579*
|
0.604**
|
0.690**
|
0.237
|
|
MAP (mm)
|
0.261
|
0.074
|
–0.103
|
–0.671**
|
–0.250
|
–0.528*
|
–0.555*
|
–0.307
|
|
GSP (mm)
|
0.244
|
0.085
|
–0.110
|
–0.689**
|
–0.269
|
–0.548*
|
–0.575*
|
–0.321
|
|
ARP (mm)
|
0.348
|
0.195
|
–0.064
|
–0.640**
|
–0.236
|
–0.503*
|
–0.495*
|
–0.365
|
|
MAT (??C)
|
–0.446
|
–0.803**
|
–0.421
|
0.534*
|
0.735**
|
0.668**
|
0.495*
|
0.860**
|
|
GST (??C)
|
–0.476*
|
–0.863*
|
–0.290
|
0.512*
|
0.672**
|
0.625**
|
0.424
|
0.880**
|
|
ART (??C)
|
0.338
|
0.503*
|
0.529*
|
–0.426
|
–0.626**
|
–0.551*
|
–0.465
|
–0.597**
|
*P <
0.05; **P < 0.01.
According
to the ANOVA GLM analysis (Table 6), the genetic variation driven by the
climate (temperature, precipitation, evapotranspiration) of the original
collection sites had significant effects on leaf anatomical structures of C.
mongholica, with between 34.09% and 81.43% of the variation explained. The
variation explained for LEC was the smallest (34.09%), and that for Tight
was the largest (81.43%). Among the climate indices of the collection sites, MAT
had a significant effect on all eight leaf anatomical indices, with very highly
significant effects on UEC, LT, UPT, LPT, PT,
and Tight (P < 0.001) and between 21.69% and 71.89% of the
variation explained. The PE significantly affected UEC, LT,
UPT, LPT, and PT (P < 0.05), with between 5.48%
and 14.34% of the variation explained. The MAP significantly affected LT, UPT, LPT, PT,
and Tight, with between 4.78% and 45.93% of the variation explained. The
MAP had very highly significant
effects on LT, UPT,
and PT (P < 0.001). The Alt had significant effects on LT, UPT, LPT, and PT,
with between 8.33% and 16.76% of the variation explained.
Table 6 Summary of general linear
models for the effects of climate variations (MAP, MAT, PE,
Alt, Site) at different collection sites on leaf anatomical structures
of Caryopteris mongholica
|
Climate factor
|
Cuticle
|
UEC
|
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
|
MAP
|
1
|
0.015
|
0.015
|
2.53
|
10.13
|
0.147
|
1
|
3 ?? 10–4
|
3 ?? 10–4
|
0.07
|
0.12
|
0.804
|
|
MAT
|
1
|
0.031
|
0.031
|
5.18
|
20.77
|
0.049*
|
1
|
0.204
|
0.204
|
34.84
|
65.82
|
2.3 ?? 10–4***
|
|
PE
|
1
|
0.006
|
0.006
|
1.02
|
4.08
|
0.339
|
1
|
0.028
|
0.028
|
4.87
|
9.20
|
0.055*
|
|
Alt
|
1
|
0.004
|
0.004
|
0.68
|
2.74
|
0.430
|
1
|
2.783
|
2.783
|
4.76
|
0.00
|
0.999
|
|
Site
|
4
|
0.039
|
0.010
|
1.63
|
26.20
|
0.248
|
4
|
0.024
|
0.006
|
1.04
|
7.86
|
0.438
|
|
Residuals
|
9
|
0.054
|
0.006
|
NA
|
36.09
|
NA
|
9
|
0.053
|
0.006
|
NA
|
17
|
NA
|
|
Total
|
17
|
0.150
|
|
|
100
|
|
17
|
0.310
|
|
|
100
|
|
|
Climate factor
|
LEC
|
LT
|
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
|
MAP
|
1
|
0.001
|
0.001
|
0.23
|
0.98
|
0.646
|
1
|
0.012
|
0.012
|
85.50
|
31.40
|
6.8 ?? 10–6***
|
|
MAT
|
1
|
0.019
|
0.019
|
3.84
|
16.78
|
0.082*
|
1
|
0.009
|
0.009
|
59.05
|
21.69
|
3.0 ?? 10–5***
|
|
PE
|
1
|
0.019
|
0.019
|
3.74
|
16.33
|
0.085
|
1
|
0.006
|
0.006
|
39.05
|
14.34
|
1.5 ?? 10–4***
|
|
Alt
|
1
|
9 ?? 10–5
|
9 ?? 10–5
|
0.02
|
0.08
|
0.894
|
1
|
0.007
|
0.007
|
45.62
|
16.76
|
8.3 ?? 10–5***
|
|
Site
|
4
|
0.031
|
0.008
|
1.52
|
26.52
|
0.276
|
4
|
0.005
|
0.001
|
8.51
|
12.50
|
0.004**
|
|
Residuals
|
9
|
0.045
|
0.005
|
NA
|
39.30
|
NA
|
9
|
0.001
|
1?? 10–4
|
NA
|
3.31
|
NA
|
|
Total
|
17
|
0.115
|
|
|
100
|
|
17
|
0.040
|
|
|
100
|
|
|
Climate factor
|
UPT
|
LPT
|
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
|
MAP
|
1
|
0.037
|
0.037
|
93.72
|
45.93
|
4.7 ?? 10–6***
|
1
|
0.004
|
0.004
|
7.47
|
4.78
|
0.023*
|
|
MAT
|
1
|
0.020
|
0.020
|
50.18
|
24.59
|
5.8 ?? 10–5***
|
1
|
0.048
|
0.048
|
90.20
|
57.74
|
5.5 ?? 10–6***
|
|
PE
|
1
|
0.005
|
0.005
|
12.82
|
6.28
|
0.006**
|
1
|
0.005
|
0.005
|
8.56
|
5.48
|
0.017*
|
|
Alt
|
1
|
0.007
|
0.007
|
17.65
|
8.65
|
0.002**
|
1
|
0.007
|
0.007
|
13.01
|
8.33
|
0.006**
|
|
Site
|
4
|
0.008
|
0.002
|
5.17
|
10.13
|
0.019*
|
4
|
0.015
|
0.004
|
6.99
|
17.91
|
0.008**
|
|
Residuals
|
9
|
0.004
|
4 ?? 10–4
|
NA
|
4.41
|
NA
|
9
|
0.005
|
5 ?? 10–4
|
NA
|
5.76
|
NA
|
|
Total
|
17
|
0.081
|
|
|
100
|
|
17
|
0.084
|
|
|
100
|
|
|
Climate factor
|
PT
|
Tight
|
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
df
|
Sum Sq
|
Mean Sq
|
F value
|
%SS
|
Sig.
|
|
MAP
|
1
|
0.019
|
0.019
|
208.39
|
27.38
|
1.6 ?? 10–7***
|
1
|
8 ?? 10–4
|
8 ?? 10–4
|
6.81
|
8.93
|
0.028*
|
|
MAT
|
1
|
0.031
|
0.031
|
337.66
|
44.37
|
1.9 ?? 10–8***
|
1
|
0.007
|
0.007
|
54.80
|
71.89
|
4.1 ?? 10–5***
|
|
PE
|
1
|
0.005
|
0.005
|
51.63
|
6.78
|
5.2 ?? 10–5***
|
1
|
6 ?? 10–5
|
6 ?? 10–5
|
0.46
|
0.61
|
0.514
|
|
Alt
|
1
|
0.007
|
0.007
|
73.49
|
9.66
|
1.3 ?? 10–5***
|
1
|
1 ?? 10–5
|
1 ?? 10–5
|
0.09
|
0.12
|
0.772
|
|
Site
|
4
|
0.007
|
0.002
|
20.20
|
10.62
|
1.6 ?? 10–4***
|
4
|
6 ?? 10–4
|
2 ?? 10–4
|
1.27
|
6.65
|
0.097*
|
|
Residuals
|
9
|
8 ?? 10–4
|
9 ?? 10–5
|
NA
|
1.18
|
NA
|
9
|
0.001
|
1 ?? 10–4
|
NA
|
11.81
|
NA
|
|
Total
|
17
|
0.070
|
|
|
100
|
|
17
|
0.009
|
|
|
100
|
|
Note:
* P < 0.05; * P < 0.01; *** P < 0.001.
5 Discussion
and Conclusion
Leaf anatomical
structures of C. mongholica from seven collection sites from 2006 to
2017 in north and northwest China in daily were measured, and the significant
environmental factors that caused differences in the structures were
determined. A common garden experiment was used to minimize the influence of
environmental gradients of the seven collection sites. Leaf anatomical structures were subject to multiple
comparisons and correlation analyses, and a general linear model was used to
identify the significant climate indices driving changes in leaf structure. The
genetic variation driven by climate was one of the major factors that caused
the differences in leaf anatomical structures from the different collection
sites. This dataset shows local adaptation in leaf structures and therefore
will help to understand the adaptation mechanisms of plants in different
regions in responding to global change. In addition, the dataset can provide basic
data for research on global change and germplasm resource protection.
Author Contributions
Liu, C., Xia,
X. L., Yin, W. L., and Ji, R. X. were responsible for the overall design and development
of the dataset. Yu, X., Chang, Y., Shen, C., Bai, X. Q., and Ji, R. X. collected
plant materials. Ji, R. X. processed leaf-related data and performed data
validation. Liu, C. designed the model and algorithm. Ji, R. X. wrote the data
paper.
Conflicts of
Interest
The
authors declare no conflicts of interest.
References
[1]
Smith, W. K., Vogelmann, T. C.,
DeLucia, E. H., et al. Leaf form and
photosynthesis [J]. BioScience, 1997, 47: 785–793.
[2]
Barboni,
D., Harrison, S. P., Bartlein, P. J., et al. Relationships between plant traits and climate in the mediterranean
region: a pollen data analysis [J]. Journal of Vegetation
Science, 2004, 15: 635–646.
[3]
Pigliucci, M. Phenotypic
plasticity and evolution by genetic assimilation [J]. Journal
of Experimental Biology, 2006, 209: 2362–2367.
[4]
Hu,
M. Y., Zhang, L., Luo, T. X., Shen, W. Variations in leaf functional traits of Stipa purpurea along a rainfall gradient in Xizang, China [J]. Chinese Journal of Plant Ecology, 2012,
36: 136–143.
[5]
Wright, I. J., Reich, P. B.,
Westoby, M. Strategy shifts in leaf physiology, structure and nutrient content
between species of high- and low-rainfall and high- and low-nutrient habitats [J]. Functional Ecology, 2001, 15: 423–434.
[6]
Chartzoulakis, K., Patakas, A.,
Kofidis, G., et al. Water stress
affects leaf anatomy, gas exchange, water relations and growth of two avocado
cultivars [J]. Scientia Horticulturae, 2002, 95:
39–50.
[7]
Zhong, Y. M., Dong, F. Y.,
Wang, W. J., et al. Anatomical
characteristics and adaptability plasticity of Populus euphratica in
different habitats [J]. Journal of Beijing
Forestry University, 2017, 39:
53–61.
[8]
Yu,
H. Y., Hu, X. Y., He, C. X., et al. Differential response of water stress on leaf
morphological anatomical structures of varied provenances Xanthocera sorbifolium [J].
Journal of Beijing Forestry University, 2019, 41(1): 57–63.
[9]
Wang, X. Q., Wang, C. K., Han,
Y. Effects of tree species on soil organic carbon density: a common garden experiment
of five temperate tree species [J]. Chinese Journal of Plant Ecology, 2015, 39: 1033–1043.
[10]
Chen, J. J., Yu, H., Xu, C. B.,
et al. Effects of provenance and
common garden environment on leaf functional traits of Machilus pauhoi
seedlings [J]. Chinese Journal of Applied and
Environmental Biology, 2019, 25:
648–654.
[11]
de Villemereuil, P., Gaggiotti,
O. E., Mouterde, M., et al. Common
garden experiments in the genomic era: New perspectives and opportunities [J]. Heredity, 2016, 116: 249–254.
[12]
Ji, R. X., Yu, X., Chang Y., et al. Leaf anatomical structure and
geographic environment data of Caryopteris
Mongholica from 7 provenances [J/DB/OL]. Digital Journal of Global
Change Data Repository, 2021. https://doi.org/10.3974/geodb.2021.01.03.V1.
[13]
Ji, R. X., Yu,
X., Chang Y., et al. Geographical
provenance variation of leaf anatomical structure of Caryopteris mongholica and
its significance in response to environmental changes [J]. Chinese Journal
of Plant Ecology, 2020, 44: 277-286.
[14]
GCdataPR
Editorial Office. GCdataPR Data Sharing Policy [OL]. https://doi.org/10.3974/dp.policy.2014.05 (Updated 2017)
[15]
Feng, Y. Q., Qin, L., Ma, H. P.
The discussion on efficiency and quality of paraffin section in plant microscopy
technique course [J]. Experimental Technology and
Management, 2008, 25: 160–162.
[16]
de Lima Silva, A., da Silva
Alves, M. V., Coan, A. I. Importance of anatomical leaf features for characterization
of three species of Mapania (Mapanioideae,
Cyperaceae) from the Amazon Forest,
Brazil [J]. Acta Amazonica, 2014, 44: 447–456.
[17]
Li, F. L., Bao, W. K. Responses
of the morphological and anatomical structure of the plant leaf to environmental
change [J]. Chinese Bulletin of Botany,
2005, 22: 118–127.
[18]
Hijmans, R. J., Cameron, S. E.,
Parra, J. L., et al. Very high
resolution interpolated climate surfaces for global land areas [J]. International Journal of
Climatology, 2005, 25: 1965–1978.
[19]
He, J. S., Wang, X. P., Flynn,
D. F. B., et al. Taxonomic,
phylogenetic, and environmental trade-offs between leaf productivity and
persistence [J]. Ecology, 2009, 90: 2779–2791.
[20]
Delzon, S. New insight into
leaf drought tolerance [J]. Functional Ecology,
2015, 29: 1247–1249.